
De første planter på landjorden
For 500 millioner år siden opstod de første landplanter ud fra grønalger, der levede i den fugtige jord omkring damme og kilder. Forfatterne giver her en aktuel status på, hvad forskerne ved om denne milepæl i livets historie.
Af Kaj Sand-Jensen og Jens Christian Schou
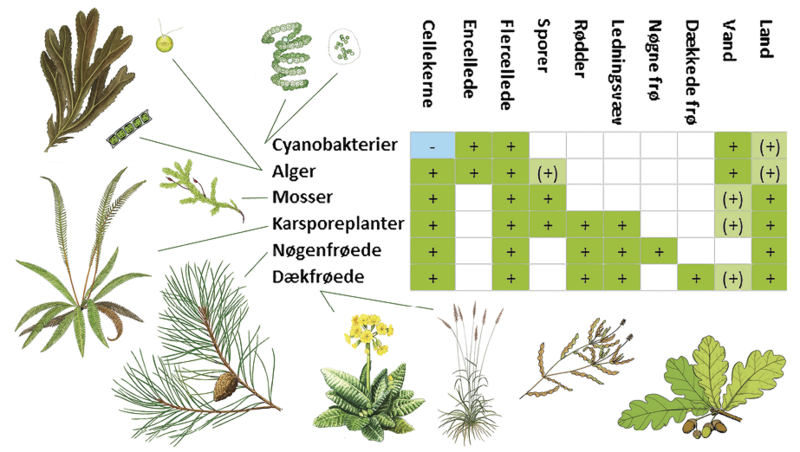
Når biologer taler om “planteliv”, omfatter det i virkeligheden tre hovedgrupper, der er i stand til at udføre fotosyntese, nemlig cyanobakterier, alger og landplanter. Cyanobakterierne er uden cellekerne, og de opstod allerede for omkring 3.500 millioner år siden. Algerne er omkring 1.800 millioner år gamle. De har cellekerne, og deres grønkorn (kloroplaster) stammer fra symbiose med cyanobakterier.
Landplanterne er det seneste skud på stammen. De opstod for blot 500 millioner år siden fra ferskvandsgrønalger. Landplanterne er flercellede og beskytter det befrugtede æg (embryoet), derfor betegnes de også embryofyter.
Vi kan således pege på tre afgørende begivenheder i plantelivets udviklingshistorie, da de tre grupper opstod første gang. Men her fokuserer vi alene på den seneste begivenhed: landplanternes opståen. Den indledte nemlig den eksplosive udvikling af mosser, karsporeplanter og de senere frøplanter, som skabte landjordens mange forskellige økosystemer og de utallige mikroorganismer, svampe og dyr, der fulgte i hælene på planterne. Under hele forløbet byggede evolutionen videre på de stoffer, strukturer og processer, som tidligere organismer havde udviklet.
Koblingsalger som landplanternes ophav
Længe før de første landplanter opstod, levede der for 2.700 millioner år siden bakteriesamfund med fotosyntetiske cyanobakterier på fugtige steder på land. De opbyggede organiske stoffer, som man i nutiden har påvist i de urgamle aflejringer.
Organiske stoffer i jorden holder på fugtigheden og frigør opløste næringsstoffer ved bakteriers omsætning, så fotosyntetiske organismer kan vokse. Da grønalger opstod for mindst 1.000-1.200 millioner år siden, kunne de derfor også leve i fugtig jord sammen med de forskellige svampe og dyr, som kom til.
I dag lever både encellede, flercellede tråde og kolonier blandt cyanobakterier og grønalger på lavt vand i søer, damme og kilder og tillige på den omgivende fugtige jord. Når jorden tørrer ud, bliver organismerne inaktive, men de liver op, når de igen får vand.
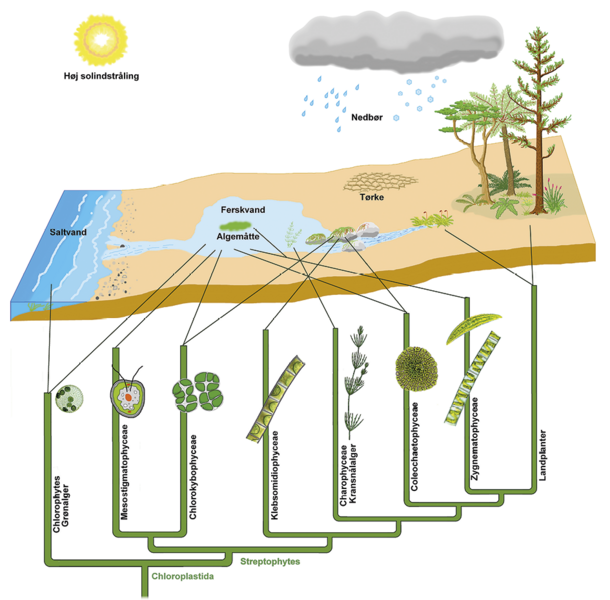
Nogle tænker måske, at de grønalger, som var forløbere for landplanterne, levede i havet, fordi havet er så stort og dækker 70 % af klodens overflade, mens ferskvandsmiljøer blot dækker omkring 1 %. Men vi ved, at landplanters ophav var grønalger fra ferskvand. Ferskvand rummer flere udviklingslinjer blandt grønalger, havet derimod blot en enkelt.
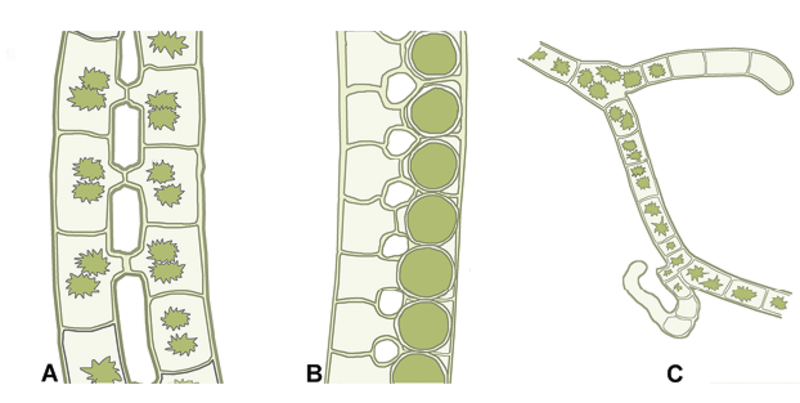
Endvidere udsprang de første egentlige landplanter af en særlig udviklingslinje af ferskvands-grønalger, som vokser på bunden af søer, damme, vandløb og i fugtig jord, men ikke findes i havet. Deres navn er koblingsalger (Zygnematophyceae).
I overgangsmiljøet fra ferskvand til fugtig jord fandtes stamformerne til de første landplanter. Stamformerne kan have haft mere komplekse væv end de nulevende koblingsalger, som således enten er mere oprindelige eller repræsenterer reducerede former. Målt på kompleksitet og størrelse ligner kransnålalger, som er en søsterlinje til koblingsalgerne, mosser og karsporeplanter mere end de nulevende koblingsalger. Kransnålalger har en hovedakse og sideakser og bliver op til 80 cm høje. Imidlertid afslører DNA og cellernes kemi og processer, at koblingsalgerne er de nærmest beslægtede med landplanter.
Fra ferskvand til landjorden
Der eksisterer en god økologisk begrundelse for ferskvands betydning som ophav til landplanter frem for havet. Kontakten mellem ferskvand og landjorden er meget mere vidtstrakt og overgangen mere gradvis end mellem havet og landjorden. Miljøskiftet fra ferskvand til landjorden sker over uforstyrret fugtig jord med ret uændrede kemiske forhold, uanset om den i perioder er tør eller sjaskvåd efter regn. Samtidigt findes dette overgangsmiljø mange forskellige steder, hvilket fremmer artsdannelsen.
Derimod er miljøskiftet brutalt både fysisk og kemisk mellem havet og landjorden. Havet har store bølger med stor fysisk kraft, og både stranden og den bølgebeskyttede kyst oplever store skift i saltholdighed. Når havvand fordamper fra saltholdig jord, omdannes jordvandet til en koncentreret saltopløsning, men efter en større regnbyge opstår en stærkt fortyndet saltopløsning. De voldsomme ændringer i saltholdigheden skaber svære vilkår for cellers overlevelse, som også i dag begrænser antallet af plantearter på kystnære jorde.
Ved bredden af damme og små søer, og hvor kildevand pibler frem af jorden, er overgangen mellem vand og fugtig jord derimod stilfærdig. Her vokser grønne koblingsalger, som har den værktøjskasse i form af gener og strukturer, som livet på land kræver.
Beskyttelse mod udtørring og UV-lys
Landjorden var alligevel på mange måder et ugæstfrit miljø for planter og deres forløbere på grund af risikoen for udtørring og frost samt skader som følge af høj solindstråling og UV-lys. Organismerne måtte beskytte og tilpasse sig til de risici, som livet på land og eksponering til luften indebar. Udtørring af cellerne kunne de forhindre ved at vokse i umiddelbar nærhed af vand. Men de måtte også holde på vandet i cellerne med overfladestrukturer, der nedsætter fordampningen.
Evnen til at overleve udtørring findes hos koblingsalger og deres efterkommere blandt mosser. De overlever udtørring ved at reducere stofskiftet til et minimum, og sparer energi til de igen får vand. Gener mod udtørringsstress kan angiveligt være overført fra jordbakterier til grønalger, hvorefter generne senere blev tilpasset livet hos landplanter. Landplanter har måske også fra begyndelsen fået hjælp ved at samarbejde med svampe, som det sker i dag for de fleste landplanter.
I en såkaldt mykorrhiza-symbiose leverer planten ofte kulhydrater fra sin fotosyntese til svampen, hvis tætte netværk af tynde levende tråde (hyfer) i jorden til gengæld optager vand og mineraler og sender dem videre til planten. En sådan symbiose er påvist hos både fossiler og nulevende arter blandt mosser og karsporeplanter.
Beskyttelse mod høj solindstråling og UV-lys på land var en anden stor udfordring for mosser på land. Mossernes forløbere blandt koblingsalger kan beskytte sig ved at indeholde UV-absorberende organiske stoffer. Vokser cellerne tæt sammen i kolonier eller i væv med mange celler, skygger de også gensidigt for hinanden og reducerer lyset og risikoen for skader på den enkelte celle.
Mossers og karsporeplanters stamformer
Som nævnt er koblingsalgerne den nærmeste søstergruppe til landplanterne, hvilket afsløres af særligt store ligheder i DNA-sekvenser, faste cellevægge og deres kemi samt mikroskopiske rørforbindelser mellem naboceller. Faste cellevægge af cellulose, hemicellulose og lignin er vigtige for at forhindre, at cellerne falder sammen ved udtørring og frost, hvilket er særlige udfordringer på land.
Koblingsalger indeholder klorofyl a og b og har stivelseskorn til oplagring af sukkerstoffer ligesom deres efterkommere blandt landplanterne. De danner deres eget “solcreme” ved at koble mange carbonringe af stoffet fenol sammen til farvede stoffer (polyfenoler). Sådanne stoffer findes overalt i planteverdenen, eksempelvis rigeligt i vindruer, der farver rødvinen. Polyfenoler er vigtige antioxidanter hos planterne og i vores plantekost, der beskytter cellerne mod skader.
Endvidere er koblingsalgernes sporer, i lighed med sporer hos mosser og karsporeplanter, beklædt med stoffet sporopollenin. Det er en kompleks polymer med blandt andet langkædede fedtsyrer og fenoler, som danner et meget fast, ydre lag, der beskytter mod udtørring, UV-lys og bakteriers og svampes nedbrydning og således sikrer sporerne langvarig overlevelse.
DNA-teknologi og landplanters evolution
DNA-teknologi og landplanters evolution
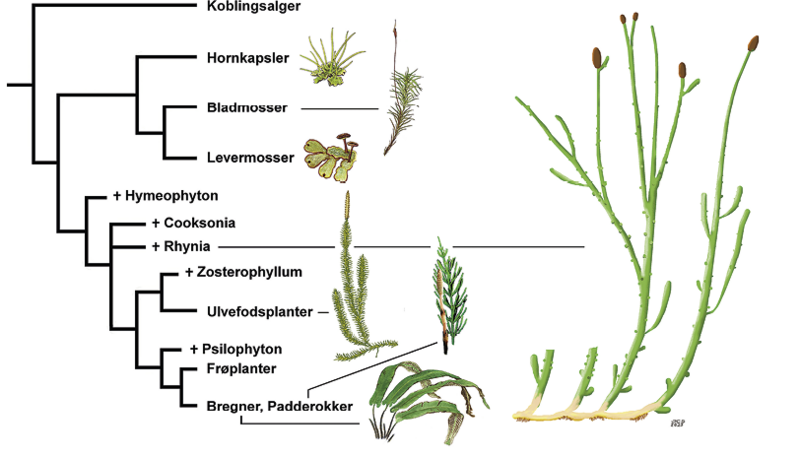
Hurtigere og dybere analyser af DNA-sekvenser hos stadigt flere arter har øget klarheden over tidspunktet for landplanternes opståen, mens der eksisterer forskellige vurderinger af de indbyrdes relationer mellem mossers og karsporeplanters evolution. I et DNA-studium fra 2018 benytter Morris og kolleger syv ligestillede modeller for evolutionen af de tre mostyper (levermosser, bladmosser og hornkapsler) og karsporeplanter. I én model udvikler karsporeplanter og mosser sig fra en fælles tidligere stamform, men i andre modeller udvikler karsporeplanter sig skiftevis fra én af de tre mostyper, hvis indbyrdes relationer også varierer, uden at den ene model er mere sandsynlig end de andre.
I et nyere studium fra 2022 af Harris og kolleger udgår både mosser og karsporeplanter fra en tidlig fælles landplante, som anses at være ganske kompleks med både simpelt ledningsvæv for vand og spalteåbninger. Ifølge denne analyse har mosserne tabt kompleksitet og gener for ledningsvæv og spalteåbninger under evolutionen og tilpasset sig en mere marginal niche, mens ulvefødder, bregner og padderokker både har tabt gener og undergået genfordobling og yderligere udvikling også i højde. Med tiden opstod frøplanter (nåletræer og blomsterplanter). Mens mosser sædvanligvis er lavere end 5 centimeter om end enkelte arter af Jomfruhår bliver 20-60 centimeter høje, udviklede træagtige karsporeplanter effektiv afstivning i 10-50 meter høje stammer, og nulevende nåletræer og løvtræer kan overstige 100 meter.
De forskellige DNA-studier er dog enige om, at landplanter indledte deres æra for 500 millioner år siden. Vurderingen bygger på det molekylære ur, som angiver den hastighed, hvormed DNA-sekvenser ændrer sig i konservative gener, der antages at være uafhængige af naturlig udvælgelse (selektion). Så gamle fossiler har man aldrig fundet. De tidligste fossiler af små karsporeplanter var indtil for nylig 430 millioner år gamle, men er nu rykket til 458 millioner år før nu i et fund fra Kina, mens sandsynlige mossporer er 470 millioner år gamle. Mange fossilforskere stiller sig skeptiske til de “hurtige” DNA-studier og understreger med rette betydningen af fossiler for sikker verifikation af evolutionshistorien. Desværre er chancen for at finde meget gamle fossiler af let nedbrydelige planter fra fugtige steder meget lav, så det er som forventet, at det molekylære ur peger længere tilbage i tiden end fossilerne.
Figuren viser et stamtræ for mosser og karsporeplanters, inklusive de uddøde karsporeplanter Hymenophyton, Cooksonia og Rhynia, som knyttes til ulvefødder og senere bregner og padderokker. På dette stamtræ har mosser og karsporeplanter (ulvefodsplanter og bregner inklusive padderokker) en parallel udvikling fra en fælles stamfader. Endvidere står levermosser og bladmosser tættere på hinanden end hornkapsler.
Planternes “hud”
Evnen til at overleve udtørring findes fortsat hos nulevende koblingsalger og deres efterkommere. Koblingsalgerne har endvidere gener for dannelse af stofferne kutin og voks, som indgår i den tynde hudagtige hinde (kutikula), der både beskytter landplanternes overflader mod udtørring og mod, at fjendtlige bakterier og svampe trænger ind udefra. Kutikulaen har vekslende tykkelse og sammensætning hos forskellige landplanter.
Både kutin og voks er langkædede fedtstoffer, som for voksens vedkommende er bundet sammen med en alkohol til det fedtagtige lag, som på landplanters blade får vanddråber til at glide af og dermed trække støv, bakterier og svampesporer med sig. Voksen er vandskyende, og hos landplanter beskytter den mod, at løvet dækkes af vand, som forhindrer optagelse af CO2 til fotosyntesen.
Hos de fleste mosser er kutikulaen tynd og nedsætter ikke fordampningen særlig meget. Mossers grønne blade eller flade løv er blot ét eller to cellelag tykt, og de mangler spalteåbninger – mikroskopiske små regulerbare porer mod atmosfæren – som karsporeplanter og frøplanter har. Derfor udveksler mosserne CO2 og oxygen ved deres fotosyntese og respiration gennem hele overfladen. Var den tynde kutikula ikke permeabel for vanddamp, ville den samtidigt standse passagen af CO2 og oxygen, og planten ville dø. Prisen for permeable mosoverflader er, at mosser ret hurtigt tørrer ud og bliver inaktive i tør luft.
Ledningsstrenge og spalteåbninger
Der var få steder, hvor de første landplanter blandt mosser og karsporeplanter kunne leve og udvikle sig, fordi de krævede en stabil vandforsyning. Mosser mangler ledningsstrenge til at transportere vand mellem fjerne dele af planten og reducere fordampningen effektivt. Tidlige karsporeplanter havde derimod simple ledningsstrenge.
Karsporeplanters grønne løv er flere cellelag tykt og har mikroskopiske spalteåbninger, der kan åbne og lukke og derved tillade eller stoppe for passage af CO2, oxygen og vanddamp.
Denne regulering har muligvis været en forudsætning for, at ledningsstrengene kunne udvikles. Når vandindholdet er højt, er spalteåbningerne åbne, CO2 passerer gennem dem og forbruges ved fotosyntesen og vanddamp og det dannede oxygen passerer ud i luften. Lukker spalteåbninger ikke ved vandmangel, risikerer planten for stor fordampning og tillige dannelse af luftbobler i ledningsstrengene, som stopper vandtransporten mere permanent, og planten visner.
De velregulerede spalteåbninger betyder samtidigt, at bladenes kutikula kunne videreudvikles til at reducere fordampningen meget effektivt, da passagen af CO2, ilt og tab af vanddamp blev koncentreret til spalteåbningerne. Planten kunne bedre holde hus med det tilgængelige vand og vokse mere for en given mængde vand.

Fra lave vækster til de første skove
De tidlige mosser var alle små og voksede tæt sammen på permanent fugtige steder. Tidlige ulvefodsplanter var også små og voksede de samme steder. Den uddøde Rhynia blev blot 6 centimeter høj og var alene fæstnet i jorden med tynde tråde i lighed med mosser og visse grønalger. Men senere udviklede ulvefodsplanter egentlige flercellede rødder, der var mere effektive til at trække vand og mineraler op fra jorden. Da fordampningen gennem bladoverfladen tillige var lav, kunne ulvefødder kolonisere tørrere jorde, og landjordens vegetation bredte sig.
Med tiden blev nogle arter til 30-50 meter høje ulvefodstræer. De indgik i Kultidens skove for 360-300 millioner år siden, som sammen med bregner og padderokker bidrog til opbygning af de store kulforekomster i våde sumpe, som vi senere har udnyttet.
En tidligere ulvefod Protolepidodendropsis pulchra med 2-4 meter høje stammer voksede i tropiske sumpe for 360-380 millioner år siden på steder, der senere blev til Svalbard.
Bregner er en artsrig gruppe af karsporeplanter, der også udgør et vigtigt indslag i Kultidens sumpskove i form af træbregner sammen med træagtige padderokker. Både bregner og padderokker dominerede fortsat landvegetationen i de efterfølgende årmillioner og var vigtig planteføde for de gigantiske dinosaurer.


Mosser og karsporeplanter i nutiden
Mossers og karsporeplanters evolution har aldrig stået stille, og de nulevende arter af mosser er således ikke overlevende relikter af de tidligste arter. På mosgrenen” i plantelivets stamtræ fraspaltes først de artsfattige hornkapsler og senere deles mosgrenen i de artsrige levermosser og bladmosser. I dag omfatter mosserne 8.000 navngivne arter af levermosser, 12.000 arter blandt bladmosser og blot 150 hornkapsler.
De fleste arter blandt nutidens levermosser vokser under fugtige forhold i de tropiske regnskove. I køligt klima danner de sjældent større selvstændige bestande. De er gennemgående meget simplere bygget end bladmosserne. Regnskoven rummer også de fleste bladmosser, men de danner tillige tætte samfund i nåleskovene herhjemme og længere mod nord. De udvikler også selvstændige samfund på fjeldheder, tundraer og på ny jord efterladt af gletsjere og vulkaner og baner vejen for blomsterplanterne.
I karsporegrenen er de uddøde Cooksonia og Rhynia knyttet til ulvefodsplanter. De øvrige karsporeplanter bregner og padderokker kom til senere.
I nutiden lever 1200 urteagtige arter af ulvefodsplanter i marginale miljøer og padderokker rummer blot 14 arter i en enkelt slægt. Bregner rummer derimod træbregner og mere end 10.000 arter, langt de fleste i tropiske skove. Blandt alle tre grupper er de fleste nye arter opstået i de seneste 66 millioner år efter Kridttiden. Det minder os om, at arterne til stadighed må tilpasse sig ændrede miljøforhold og nye arter skal opstå, for at plantegrupperne ikke skal uddø. ♦


Kaj Sand-Jensen er professor i ferskvandsøkologi ved Biologisk Institut, Københavns Universitet og forfatter til bøger om Danmarks natur og biodiversitet samt lærebøger om økologi.
ksandjensen@bio.ku.dk
Jens Christian Schou er botaniker, forfatter og pensioneret lærer.
jcschou@gmail.com