
Flagellater – havets mikroskopiske prædatorer
De fundamentale livsfunktioner – at æde og overleve – er hos alle organismer i konflikt med hinanden. Denne konflikt hos såkaldte flagellater er en nøgle til at forstå de mikrobielle økosystemers funktion i havet og dermed havets rolle i det globale kulstofregnskab.
Af Thomas Kiørboe, professor, DTU-Aqua
Vi er alle rundet af en flagellat: sædcellen der med sin piskende flagel svømmer mod ægget minder os om vores marine urform. Den encellede, fritlevende flagellat opstod for mere end 1 milliard år siden og gav ophav til alle nulevende eukaryoter (det vil sige alt liv, der ikke er bakterier og vira). Flagellater findes stadig i alle vandige miljøer og i mange forskellige varianter. Dyr og planter er alle små sidegrene på det centrale eukaryote stamtræ, og langt den største evolutionære diversitet findes blandt flagellater. Faktisk findes flagellater i alle stamtræets hovedgrene, og det er således den mest diverse gruppe organismer på jorden.
Fritlevende flagellater spiller en central rolle for reguleringen af havets mikrobiom, det vil sige de mikrobielle økosystemer, der står for hovedparten af den biologiske carbonomsætning i havet. Flagellater lever af bakterier, og de fleste er tillige i stand til at lave fotosyntese og dermed optage CO2. De ernærer sig altså både som dyr og planter. Flagellaterne er de største konsumenter af bakterier i havet og vigtige primærproducenter af organisk carbon. De spiller altså en central rolle for havets bidrag til det globale carbonregnskab og dermed for vores klima.
Modellering af økosystemer
For at vurdere havets rolle i det globale carbonregnskab – og hvordan denne rolle påvirkes af blandt andet klimaændringer – bruger vi modeller af marine økosystemer.
Men modellering kræver forenkling: Det er i praksis ikke muligt at beskrive alle arter, og hvordan de vekselvirker med hinanden og det omgivende miljø. Derfor slår man ofte arter, der ligner hinanden, sammen i funktionelle grupper, som man behandler som var det én art. Således beskriver man i mange matematiske modeller flagellater som én funktionel gruppe: De er alle små, encellede og udstyret med nogle få flageller. Derved ser man bort fra, at de indbyrdes er meget forskellige:
De varierer kolossalt i størrelse (de største vejer mere end en million gange mere end de mindste); de varierer i form – flade, runde, aflange og med diverse udvækster; deres flageller kan variere i antal og være forsynet med hår og sejl, og deres slagmønstre kan være meget forskellige. At behandle flagellater som èn gruppe svarer omtrent til at behandle alle dyr og alle planter som hver én gruppe i beskrivelsen af økosystemers sammensætning og funktion.
Derfor bruger moderne “træk-baserede” modeller et andet forenklingsprincip. I stedet for at slå arter sammen i funktionelle grupper, fokuserer man på individuelle organismers vigtigste egenskaber (træk) på tværs af arter. Der skal ikke mange egenskaber til for at få en god beskrivelse af en organismes vigtigste livsfunktioner, at æde og overleve. Ved at kvantificere de indbyggede modsætninger mellem de fundamentale livsfunktioner kan man konstruere simple modeller, hvor fødenettets komplicerede struktur og funktion opstår som et resultat af interaktioner mellem individer og miljø. Modelbyggeren skal ikke selv specificere “hvem der æder hvem”. Spørgsmålet er, om vi kan kvantificere modsætningerne – trade-offs – mellem at æde og overleve for flagellater i modelleringen af marine mikrobielle fødenet?
Flagellaternes modsætningsfyldte liv
Flagellater bruger flagellen til at skabe en vandfødestrøm, hvorfra den kan høste bakterier. Men når flagellaten forstyrrer det omgivende vand med sin fødestrøm, tiltrækker den samtidig sine fjender, og der bliver derfor en konflikt (trade-off) mellem at æde og selv blive ædt.
I flagellatens mikromiljø fremstår vand så sejt som sirup. Det vil sige at den svømmende flagellat slæber en tyk kappe af sejt vand med sig, og hvis den nærmer sig et bytte, vil dette alt andet lige blive skubbet væk. Der skal altså ganske store kræfter til for at skabe en effektiv fødestrøm, der trækker byttet til sig.
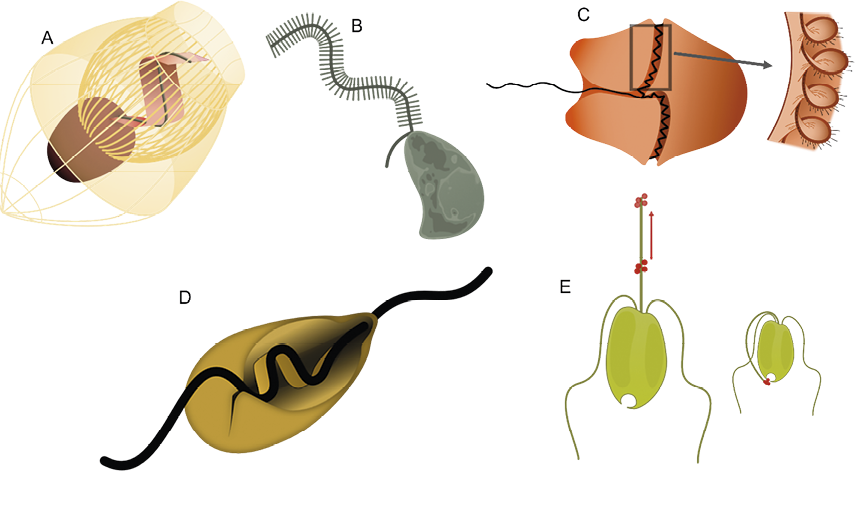
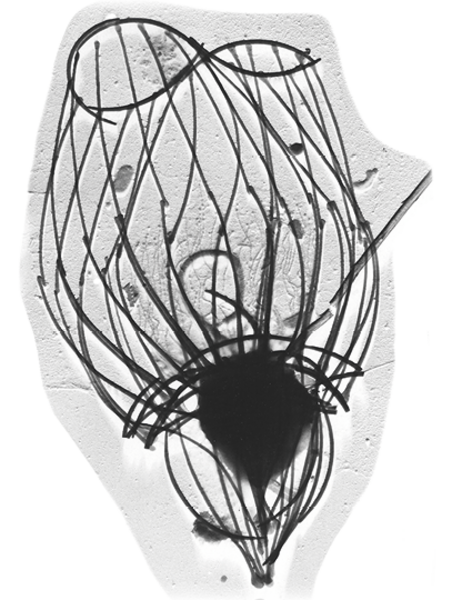
Figur 1. Flagellater har forskellige tilpasninger til at drive en effektiv fødestrøm.
A. Kraveflagellat: flagellen er omgivet af en “filterkrave”, der består af tætsiddende filamenter. Flagellen driver en bred hinde, der fungerer som en effektiv pumpe. Mange kraveflagellater er, som denne, desuden omgivet af et åbent eksoskelet, der består af sammenhæftede stave.
B. Flagellat med stive hår på den ene flagel. Hårene 10-dobler den kraft, flagellen producerer og vender kraftens retning, så fødestrømmen kommer ind mod cellen. C. Dinoflagellat (furealge) med èn nøgen flagel og èn flagel, der indpakket i en hinde løber i en fure rundt om cellen og producerer en kolossal kraft, der skaber en meget effektiv fødestrøm.
D. Excavat flagellat med to flageller, hvoraf den ene driver en bølgende hinde på cellens underside.
E. Haptofyt udstyret med to flageller samt en “snabel”, hvorpå bakterier opsamles og føres til flagellatens mund. Tegningerne er ikke til skala – de afbillede flagellater måler mellem 4 og 30 µm i diameter.
Hvordan skaber en flagellat en kraftig fødestrøm?
Den arketypiske flagellat, der drives frem af en enkelt flagel, finder vi hos den gruppe, hvortil dyrene hører. Sædcellen hos dyr er et godt eksempel; sværmerstadier hos kraveflagellaterne (vores nærmeste slægtninge blandt flagellaterne) et andet. Men den nøgne flagel kan ikke skabe en tilstrækkelig kraft til at drive en effektiv fødestrøm, og vi finder derfor en række tilpasninger hos det stadie, der æder.
Ernæringsstadiet hos kraveflagellater etablerer således en krave omkring flagellen (fig. 1A). Kraven består af tætsiddende stave og fungerer som et filter. Når flagellen slår, drives der vand gennem dette filter, og her tilbageholdes bakterier. Kun ved at antage, at flagellen er udstyret med brede hinder, har man kunnet forklare væskestrømmen gennem filtret. Det er først for nylig lykkedes at visualisere denne skrøbelige hinde.
Andre flagellater har stive hår på flagellen (fig. 1B). Men hårene stikker ud fra flagellen i slag-planet, ikke vinkelret, som man ville forestille sig mere virkningsfuldt. Og tilstedeværelsen af hår vender strømmen, så den løber modsat den retning, flagellen slår, og samtidig øges flagellens kraft med en faktor 10. Uforståeligt? På denne skala, hvor vand opfører sig mærkeligt, kan man ikke bruge sin intuition til at forstå det, man observerer. Ved hjælp af formel fysik kan vi imidlertid forklare observationen. Strømning ind mod i stedet for væk fra cellen har store fordele. Flagellaten scanner ved hjælp af flagellen den indkommende fødestrøm for bakterier og transporterer herefter opsnappet bytte til cellen, godt hjulpet af fødestrømmen. Denne effektive type flagellater syntes at være den dominerede form i havet.
Furealger (dinoflagellater) har et andet unikt arrangement (figur 1C): de har to flageller, den ene minder om sædcellens og skubber cellen fremad, mens den anden flagel løber i en fure rundt om cellen. Denne flagel ligger desuden i en hinde, som den driver i en bølgende bevægelse rundt om cellen. Endelig er den tillige udstyret med hår. Igen kan vi kun ved hjælp af formel fysik forstå, at dette system producerer en endog meget stor kraft, som dels driver cellen fremad, og dels skaber en effektiv fødestrøm, der fører vand og bytte gennem den seje vandkappe helt ind til cellens overflade.
De “excavate” flagellater (figur 1D) har ligeledes to flageller, hvor den ene, forsynet med en hinde, driver en strøm gennem en fure på cellens underside, hvor byttet indfanges. Systemet synes ikke at være særlig effektivt, og de fleste nulevende former findes enten i havbunden, eller fasthæftet på svævende partikler (“marin sne”), hvor koncentrationen af føde er høj. Excavaterne anses af nogle forskere for at være nært beslægtet med den fælles stamform for alle nulevende eukaryoter. I ur-havet var behovet for en effektiv fødestrøm mindre end i det moderne hav, fordi der var færre prædatorer.
Endelig er der “haptofytterne” (figur 1E), som er celler, der ud over to flageller er udstyret med et “haptonem”: en lang, tynd udvækst, hvor cellen opsamler bakterier og som en snabel kan føre byttet til “munden” i cellens anden ende.
Der findes mange andre typer flagellater i havet. Mange af dem kender vi kun fra det eDNA, man kan isolere i vandprøver, og mange andre kan ikke dyrkes og studeres i laboratoriet. Men vi må forvente, at de alle har tilpasninger til at producere tilstrækkeligt effektive fødestrømme.
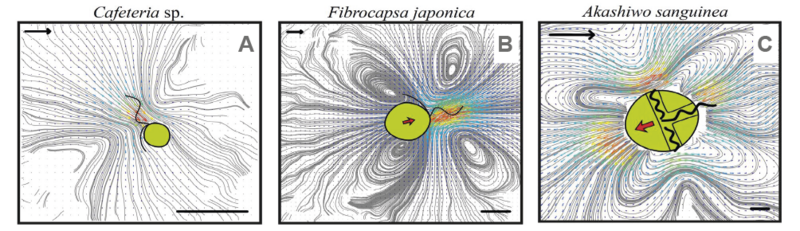
Figur 2. Fødestrømmens arkitektur varierer på karakteristisk vis mellem arterne, som alle fordeler sig i 4 forskellige generiske grupper (her er vist 3; den 4. gruppe minder i sin arkitektur om typen vist i C). Linjerne er “strømlinjer”, som er den bane en vandpartikel følger. Længdeskala i nederste hjørner er 10 µm, og flowskalapile i øverste venstre hjørne er 0,1 mm/s.
Prædations-risiko
Jo kraftigere og jo længere væk, fødestrømmen rækker, des mere bytte kan høstes, men des større er også flagellatens risiko for at blive opdaget af et rovdyr, for eksempel en vandloppe. Vandlopper er udstyret med små sansehår, der som en pick-up på en gammeldags pladespiller opsnapper bevægelser i vandet. Fødestrømmens udbredelse og arkitektur varierer på karakteristisk måde mellem forskellige typer flagellater (figur 2).
Den er relateret til, hvor og hvordan flagellen fungerer og kan forklares med simple hydrodynamiske principper. Hvis flagellaten er fasthæftet, er fødestrømmens udbredelse større, end hvis den svømmer, og hvis flagellens kraft primært virker omkring cellens “ækvator”, som hos dinoflagellater, aftager forstyrrelsen hurtigt med afstanden til cellen. Hos dinoflagellaterne kompenserer det i nogen grad for den usædvanligt store kraft og effektive fødestrøm, flagellen producerer.
Analyser af fødestrømmens udbredelse og dens effektivitet illustrerer den fundamentale konflikt mellem at æde uden selv at blive ædt: Individer, der er effektive til at samle føde, udsættes også for den største prædationsrisiko (figur 3). De hydrodynamiske modeller hjælper os til at forstå de forskelle mellem gevinst og risiko, man kan se mellem de forskellige arkitekturer.

Figur 3. Flagellater med den mest effektive fødesøgning udsættes også for den største prædationsrisiko. Skalaerne på de to akser er logaritmiske og relative: for eksempel vil en organisme med en prædationsrisiko på 100 (= 1) løbe en 10 gange så stor risiko for at blive ædt af en prædator, som en med risiko101 (= 10). Tilsvarende med fourageringseffektivitet på X-aksen.
Symbolerne refererer til de 4 principielle arkitekturer af fødestrømmen, som flagellater kan inddeles i. Den faktiske risiko og fourageringseffektivitet afhænger af, hvor mange prædatorer, og hvor meget føde, der er i det omgivende miljø. Der er karakteristiske forskelle i forholdet mellem risiko og fødesøgningseffektivitet mellem de 4 grupper.
Hvordan kan vi forenkle?
Ovenfor har jeg beskrevet noget af den store funktionelle variation, vi finder mellem forskellige grupper af flagellater. Der er mange arter, men relativt få typer, der kan karakteriseres ved fødestrømmens arkitektur på tværs af arter. Og vi kan herudfra kvantificere den fundamentale konflikt mellem at æde og overleve. Sådanne konflikter er den naturlige selektions substrat, det vil sige kilden til artsrigdom, og dermed bestemmende for økosystemers sammensætning og funktion. Ved i første omgang på nørdet vis at fokusere på forskelle mellem arter og på de underliggende mekanismer kan vi efterfølgende på meningsfuld måde forenkle og generalisere uden at skulle beskrive alle arter.
Og vi kan formulere vores indsigt ved hjælp af simple matematiske modeller, som så igen egner sig til at indgå i de globale modeller af marine mikrobielle systemer, som vi har under udvikling. Men det er en anden historie. ♦
Om forfatteren

Thomas Kiørboe er professor, dr. scient. og leder af Centre for Ocean Life ved DTU-Aqua. Han forsker i samspillet mellem plankton-organismer og det omgivende miljø.
tk@aqua.dtu.dk