
Blomsterplanterne førte biodiversiteten til usete højder
Blomsterplanternes udvikling af utallige landlevende arter siden Kridttiden skabte en makroøkologisk revolution blandt tilknyttede insekter, edderkopper, hvirveldyr og svampe, som bragte biodiversiteten til helt nye højder. Hvilke egenskaber hos blomsterplanterne har drevet denne udvikling, som Darwin kaldte et mysterium?
Af Kaj Sand-Jensen og Jens Christian Schou
Planter har en kort tidshistorie på landjorden på blot 460-500 millioner år sammenlignet med de mindst 2 milliarder år, hvor der har levet alger i ferskvand og havet. De første landplanter var mosser, der udvikledes fra ferskvandsgrønalger af typen koblingsalger.
Mosvegetationen var blot få cm høj og kun tæt på fugtige steder, da mosser mangler egentlige rødder til at optage vand fra jorden og ledningsstrenge til at føre vandet op i planten.
Den næste æra tilhørte karsporeplanterne. Først ulvefødder, for cirka 430 millioner år siden, udstyret med et simpelt ledningsvæv til vandtransport, spalteåbninger i bladene og flercellede korte rødder. Senere i Kultiden (359-299 millioner år) voksede træagtige padderokker, bregner og koglepalmer sammen med de nøgenfrøede nåletræer i skove, der dominerede landjordens vegetation helt frem til Kridttidens begyndelse (145 millioner år), hvorefter blomsterplanter overtager scenen. Og det gør de meget hurtigt og i stort artsantal ledsaget af et endnu større mylder af nye tilknyttede arter blandt specialiserede insekter, edderkopper, hvirveldyr og svampe. Darwin kaldte denne udvikling et “afskyeligt mysterium” (abominable mystery).
Siden Darwin er vi blevet meget klogere på blomsterplanternes nye bygningstræk og livsytringer samt deres slægtskab og evolution. I denne artikel forklarer vi, hvordan blomsterplanternes store genetiske fleksibilitet, højere stofproduktion og forskelligartede blomster over tid skabte grundlaget for en samlet global artsdiversitet på flere millioner arter.
Da blomsterplanterne indtog scenen
Blomsterplanterne blev for alvor talrige midt i Kridttiden for 100 millioner år siden. I de tidlige faser levede de især i ferskvand og på forstyrrede flod- og søbredder. Med tiden bredte de sig i bunden af datidens nåleskove. Først senere udviklede de egentlige træer, der udkonkurrerede nåletræerne i varme klimaer.
Midt i Kridttiden var blomsterplanternes bidrag til artsantallet i lokale floraer beskedne 5-20 %. Men ved slutningen af Kridttiden for 66 millioner år siden omfattede de mere end 80 % af floraens arter. Ved sammenligning af basesekvensen i DNA-strengene og vurdering af den hastighed, bestemte baser kan mutere over tid, har forskerne udpeget nogle få tidlige blomsterplanter, heriblandt åkander, som senere følges af de to meget artsrige enkimbladede planter med blandt andet græsser og orkidéer og tokimbladede med hele buketten af nelliker, kurvblomster, ærteblomster og roser. Udspaltningen i mange familier og slægter skete 100-50 millioner før nu, men endnu flere slægter og arter fulgte, så antallet af beskrevne blomsterplanter nu tæller over 300.000 arter.
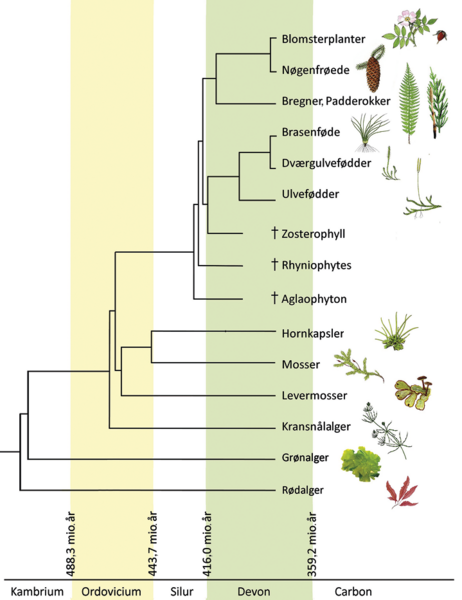
I dag er artstallet af forløberne blandt ulvefødder meget lavt, og de nøgenfrøede, hovedsageligt nåletræer omfatter blot 1100 arter. Antallet af navngivne mosser (21.925) og bregner plus padderokker (10.560) er heller ikke prangende. De er alle trængte af konkurrence om pladsen med de talrige blomsterplanter.
Måske undervurderer vi artsrigdommen blandt uddøde arter i de ovennævnte plantegrupper i fortiden. Fossilerne repræsenterer trods alt et begrænset udsnit af de tilstedeværende arter. Men flere vurderer, at artsrigdommen blandt nutidens mosser og bregner er mindst lige så høj som førhen. Er det korrekt, er landplanternes artsrigdom i dag mindst ti gange højere end i fortiden, da mosser, karsporeplanter og nøgenfrøede var enerådende.
Tilpasninger til bedre udnyttelse af CO2 og vand
Der ligger sandsynligvis både ydre faktorer og biologiske egenskaber bag blomsterplanternes overvældende artstal. Siden Kridttiden er gennemsnitstemperaturen for vandsøjlen i verdenshavene faldet fra omkring 20 til få grader, dog med markante varmeperioder undervejs.
Superkontinentet Pangea blev opdelt i to store klumper for 200 millioner år siden. I løbet af Kridttiden vandrede Sydamerika og Afrika mere fra hinanden, og et bredere sydligt Atlanterhav blev dannet. I sen Kridttid blev Australien og Antarktis adskilt og Grønland frigjorde sig fra Nordamerika. Flere kontinenter og nye bjergkæder adskilte områder med forskelligt klima og skabte muligheder for udvikling af nye arter på kontinenterne og i isolerede bestande i dale og på bjergskråninger. Blomsterplanterne har af forskellige grunde været meget bedre end karsporeplanter og nåletræer til at tilpasse sig og udvikle nye arter under de vekslende varme og kølige samt fugtige og tørre forhold.
CO2-indholdet i atmosfæren har varieret betydeligt over perioden, og det faldt fra omkring 800 ppm (dele per million) i Kridttiden til blot 280 ppm umiddelbart før industrialiseringen, hvilket stillede krav til tilpasningsevnen. Fald i atmosfærens CO2-indhold udløste ændringer i bladanatomi og biokemi i adskillige plantefamilier. Det kommer til udtryk i en ny type fotosyntese, man kalder C-4- fotosyntese, der findes hos varmeelskende planter (for eksempel majs) i tørre miljøer for at opretholde fotosyntesen ved lav tilgængelighed af CO2 og vand.
Planter med C-4-fotosyntese koncentrerer CO2 inde i bladet omkring det enzym (RUBISCO), der katalyserer indbygning af CO2 i nye kulhydrater, så processen kan forløbe meget effektivt, når spalteåbninger er åbne. Til gengæld kan planten efterfølgende lukke spalteåbningerne i længere tid og dermed mindske den samlede fordampning. C-4-fotosyntesen fungerer dermed som tilpasning til både højere temperaturer og tørre forhold.
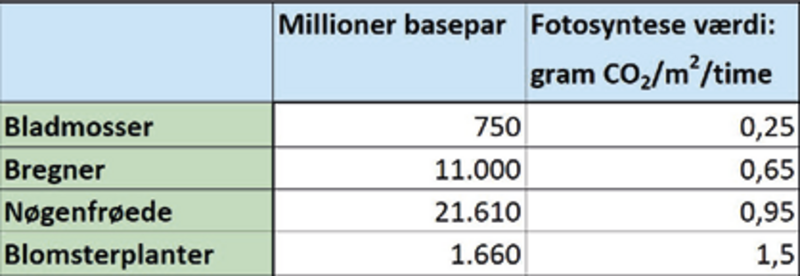
Blomsterplanterne har udviklet andre tilpasninger til at udnytte CO2 og vand bedre. De to vigtigste var større effektivitet af ledningsstrengene i vandtransporten og flere spalteåbninger med bedre kontrol over åbning og lukning, så optagelsen af CO2 til fotosyntesen i forhold til vandtabet ved fordampning optimeres. Mens karsporeplanter og nøgenfrøede planter har ledningsstrenge med tværvægge, udvikler blomsterplanterne egentlige kar, uden tværvægge og sædvanligvis større diameter, der gør dem i stand til at transportere mere vand.
Blomsterplanterne har også større tæthed af ledningsstrenge i bladene til at forsyne cellerne. Desuden kan bladcellerne hos blomsterplanterne være mindre end hos ulvefødder, bregner og nåletræer, hvorved cellernes overflade i forhold til volumen bliver større og fremmer CO2-optagelsen.
Alt i alt betyder bedre vandforsyning, mindre bladceller og flere spalteåbninger, at blomsterplanter kan opnå højere fotosyntese og tilvækst end ulvefødder, bregner og nåletræer. Den højere fotosyntese kan levere et overskud af organisk stof til mange formål. For eksempel til at danne et dybere rodnet, som kan skaffe mere vand under tørke. Eller til at etablere blomster med særlige strukturer og lokkende dufte, som kan fremme bestøvning og frøproduktion.
Udvikling af blomsten
Da blomsterplanterne udviklede blomster fik de dermed også en ny forplantningsbiologi. Det fremmede både dannelsen af nye arter og deres overlevelse. De tvekønnede blomster, der lokker især insekter med farver og dufte samt belønner med sukker i nektar og protein i pollen, sikrer blomsternes krydsbestøvning og genetiske variation.
Blomstens pollen og æg er beskyttede i adskilte stærkt omdannede støvblade (støvdragere) og frugtblade (frøanlæg). Pollen kan spredes effektivt til fremmede blomsters frøanlæg, og befrugtningen sker uden medvirken af vand. Frøet spredes og kan direkte vokse frem til en ny plante.
Ulvefødder og bregner spreder sporer, der spirer på fugtige steder til en lille grøn kim, hvor befrugtningen sker, hvorefter en ny sporedannende plante kan vokse frem. Den selvstændige kims krav på høj fugtighed udgør derfor en snæver flaskehals for disse karsporeplanters spredning og udvikling, som blomsterplanter helt undgår.
Senere ændringer i blomstens opbygning fremmede artsudviklingen og spredningen. Blomsterne udviklede sig fra en åben, radiærsymmetrisk blomst mod en mere lukket en-symmetrisk blomst, som hos ærteblomster, læbeblomster og orkidéer, der bestøves af insekter og endog specialiserede arter. Det sikrer mere præcis krydsbestøvning, når insekterne besøger netop denne ene slags blomst. På vindombruste åbne sletter har vindbestøvning omvendt været en succes hos de mange græsarter.
Fintunede samspil mellem blomsterplanter og dyr
Fintunede samspil mellem blomsterplanter og dyr

Blomsterplanter indeholder et kæmpe arsenal af attraktive og frastødende egenskaber for dyrene. Darwin var opmærksom på den gensidige fintunede afhængighed, der kan findes mellem bestemte blomsterarter og insektbestøvere og dens betydning for den gensidige artsdannelse.
Darwins orkidé (Angraecum sesquipedale) er en epifyt i Madagaskars regnskov med en 25-43 cm lang blomsterpore. I 1862 forudså Darwin, at der eksisterede et insekt med en tilsvarende lang skræddersyet sugesnabel.
Insekter med kort sugesnabel kan godt frigive pollenmasserne og få dem på sig i et forgæves forsøg på at få fat i nektaren, men kun individer med lang snabel vedbliver med at besøge og dermed bestøve blomsten lokket af nektarbelønningen. Darwins forudsigelse blev et fejret eksempel på evolutionsteorien og mulig co-evolution mellem blomsterart og insektart. Det blev bekræftet i 1903, længe efter Darwins død, da man fangede en aftensværmer med netop så lang en sugesnabel, men først i 1990’erne så man den bestøve blomsten.
Siden har man diskuteret, om blomsten og insektet gensidigt har tilpasset sig til hinanden, eller det alene er blomsten, som ved naturlig udvælgelse er blevet tilpasset til den langtungede aftensværmer, som førhen har bestøvet andre dybe blomster.
Kæmpestore forskelle i størrelse og opbygning af frø og frugter er også udviklet som tilpasning til forskellige spredningsveje ved hjælp af enten insekter, fugle, pattedyr, vind eller vand. Hundredtusinder mikrogram-lette frø spredes med vinden fra enhver blomsterkapsel hos orkidéer, stæreflokke kaster sig over kirsebær, mens ræven får frø med krogbørster i skindpelsen og spreder dem, når den lusker gennem skoven.
Eksplosion af dyr og svampe
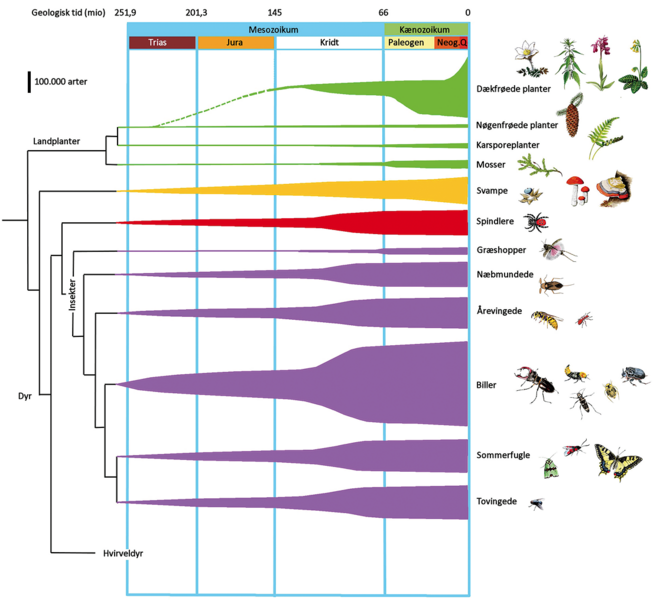
I de seneste 100 millioner år er der sket en eksplosion af landlevende dyr og svampe knyttet til blomsterplanterne. I runde tal omfatter navngivne landlevende arter op mod to millioner. Landjordens artsdiversitet er dermed cirka 5 gange større end den samlede artsdiversitet i ferskvand og havet. Langt den største hovedgruppe på land er insekterne med godt 1 million navngivne arter. Her er artstallet i de største hovedgrupper, biller (387.000), sommerfugle (158.000), fluer og myg (156.000), bier og myrer (117.000) samt tæger (104.000).
Edderkopper plus mider og svampe rummer begge omkring 100.000 arter. Den høje artsdiversitet af insekter og svampe er ikke overraskende, da adskillige specialiserede arter blandt bladlus, dværgmøl, snyltemøl og rust- og brandsvampe er knyttet til hver eneste planteart. Andre insekter flyver fra blomst til blomst og udnytter nektar og pollen, eller sidder stille, suger plantesaft og æder bladene, veddet eller rødderne. Atter andre lever af døde blade i jorden, som rovdyr eller som specialiserede parasitter.
Der er sikkert en yderligere grund til, at så mange insektarter er knyttet til blomsterplanterne. Insekterne har – i lighed med alle andre planteædere – brug for meget protein, nitrogen og fosfor, og derfor foretrækker de at æde de mest næringsrige plantedele. Blomsterplanter er rigere på protein, nitrogen og fosfor, vokser hurtigere og konsumeres også i højere grad af dyrene end ulvefødder, bregner og nøgenfrøede planter.
Blomsterplanterne har nemlig udviklet særlige evner til at kunne indbygge næringsstoffer i vævet, sædvanligvis i højere grad end karsporeplanter og nøgenfrøede. Fosfor dannes ikke biologisk, men mange blomsterplanter udskiller citronsyre og oxalsyre fra rødderne og opløser jordens kalkfosfater, så fosfat kan optages. Men endnu vigtigere lever blomsterplanter i et mutualistisk symbioseforhold (mykorrhiza) med svampe, hvis netværk af tynde hyfer afsøger store jordvolumener for fosfor, som de opløser med enzymer og transporterer til værtsplanten, der kvitterer med organisk stof fra sin høje fotosyntese til svampene. Orkidéerne har helt reduceret rodnettet og overladt det til svampepartnerne at levere vand og opløste næringssalte.
Visse blomsterplanter har den særlige egenskab at kunne fiksere luftformigt nitrogen (N2) som biologisk anvendeligt ammonium i rodknolde med bakterier. Egenskaben er udbredt hos 3000 arter i ærteblomstfamilien. De kan derfor kolonisere jomfruelig jord uden plantetilgængeligt nitrat og ammonium foran smeltende gletsjere, i floddeltaers sand og i andre nitrogenfattige jorde. Når ærteplanterne dør og nedbrydes, beriger de jorden med nitrogen og baner vejen for andre blomsterplanter.
Fordobling og slankning af genomet
Fordobling og slankning af genomet
Hybrider bringer genetisk information fra forskellige forældrearter sammen, og hybriden er sædvanligvis steril, men kan formere sig vegetativt. Hybrider kan være yderst livskraftige og have årelang overlevelse. I nogle tilfælde fordobles antallet af kromosomer i hybriden, og i disse tilfælde kan den nye plante gennemføre en reduktionsdeling (meiose), danne pollen og æg, blive krydsbestøvet og producere frø med nye egenskaber.
På trods af gentagne kromosomfordoblinger har blomsterplanterne opretholdt et lille genom ved at fjerne “unødvendige” kromosomer og DNA-kopier. Et genom rummer egentlige gener, der koder for proteiner, andet DNA, som koder for RNA, der tænder og slukker for generne, og endelig DNA, der påvirker tænd-sluk-mekanismen og dermed fænotypen. Vi ved ikke, hvor meget af genomets DNA, som er overflødigt for plantens fremtoning.
Men et lille genom kræver mindre energi og fosfor og kvælstof til DNA og det omfattende apparat af ribosomal-RNA, rRNA, messenger-RNA, mRNA, energimolekylet ATP og proteiner, som fordobler DNA’et ved celledeling. Da fosfor og nitrogen ofte begrænser arters udbredelse og vækst, klarer planter med fordoblede kromosomtal og et stort genom sig bedst på næringsrige lokaliteter, mens forældrearter bedre tåler næringsfattige betingelser.
Fordelen ved et lille “fleksibelt” genom
En stor variation i generne og deres regulering ligger bag de mange arter og livsformer blandt blomsterplanterne. Man forsker i, om denne variation kan skyldes særlige egenskaber ved genomet. På den ene side har der været mange kromosomfordoblinger efter hybriddannelse mellem to arter, som fordobler genomets størrelse og kan tilføre nye egenskaber og skabe nye arter. Spontan kromosomfordobling af genomet uden fremmede arters indblanding er også hyppig og kan eksempelvis fjerne effekten af skadelige mutationer på plantens udvikling og mindske risikoen for, at det normale gen tabes over generationer, når det er til stede i dobbelt antal. Efter kromosomfordobling slankes genomet sædvanligvis over årmillionerne til dets oprindelige størrelse.
Kromosomfordoblinger kan forbedre tilpasninger i perioder med voldsomme klimaændringer mod varmere eller koldere og tørrere forhold, og denne positive effekt er tydelig hos græsser, kurvblomster og ærteblomster. Kromosomfordoblinger sker også med øget hyppighed blandt blomsterplanterne på udfordrende kolde levesteder i bjerge og arktiske egne.
Denne fleksibilitet – at kunne fordoble DNA’et og efterfølgende fjerne det overflødige – er veludviklet hos blomsterplanterne. I hvert fald har bregner og nåletræer i gennemsnit 7 og 13 gange større genomer end blomsterplanterne, og det koster mere at producere de større genomer. Slankning af genomet mindsker forbruget af næringsstoffer, energi og tidsforbruget til genomets fordobling ved celledelinger. Halvdelen eller mere af cellens fosfor og meget nitrogen kan være bundet i DNA og ribosomernes RNA, hvilket især er kritisk for planter på næringsfattige jorde.
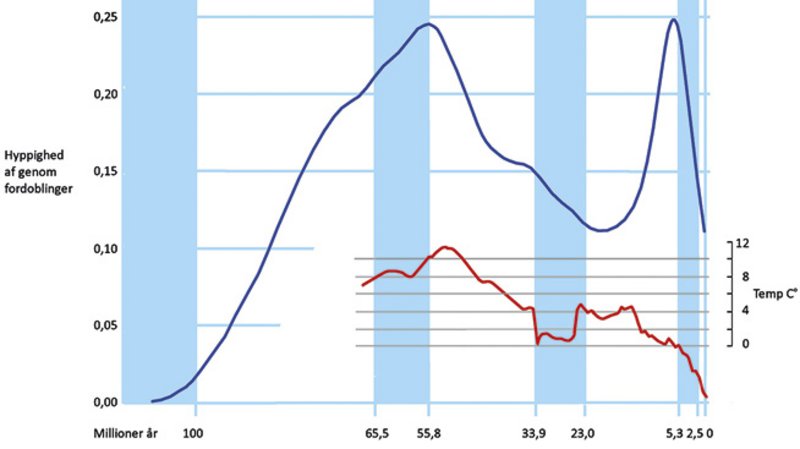
Meget af genomet er “dødvægt” (junk-DNA), som ikke er gener, der danner proteiner eller er virksom som struktur- eller genregulerende RNA i cellerne. “Dødvægten” er særlig tung i store genomer, som mange derfor mener er en blindgyde i evolutionen, som arten kun kan klemme sig ud af ved at slanke genomet.
Biodiversitetens udvikling
Mange år efter Darwin belyser nye DNA-data både blomsterplanternes evolution, slægtskab og indbygning af nye gener og deres funktion. Vi er kommet tættere på at kunne forklare de mange arter af blomsterplanter og deres tætte relationer til insekter, svampe og bakterier, som for Darwin var et mysterium.
Blomsterplanterne er som det sidste skud i landplanternes evolution i stand til at vokse mange flere steder, dække jordoverfladen bedre med en vegetation i flere lag og udvikle et tættere og dybere rodnet end karsporeplanter og nøgenfrøede. Blomsterplanterne er dermed bedre til at udnytte lys, vand og næringsstoffer og producere mere organisk stof. Deres stofproduktionen er desuden gennemgående af bedre næringskvalitet, og det er grundlaget for mange flere og mere specialiserede tilknyttede arter.
Det er sikkert også sådan, at årmillionerne i sig selv har arbejdet for øget udspecialisering af stadigt flere arter, der rækker videre end de muligheder, som den øgede planteproduktion alene skabte. Det har sandsynligvis været tilfældet i hele livets knapt 4 milliarder års udvikling. Der har dog været bump på vejen ved voldsomme geologiske hændelser, der reducerede artstallene i nogle millioner år, hvorefter tallene igen steg til nye højder.
Denne fremadskridende stigning af biodiversiteten ændrede sig, da det moderne menneske for 60.000 år siden forlod Afrika og befolkede alle egne på jorden. Og det vendte til en begyndende reduktion af biodiversiteten, da menneskets befolkningstal steg til mange milliarder og lagde beslag på store arealer til produktion af mad til at mætte de mange munde. ♦

Giftige stoffer findes i utallige planter, for eksempel opium i valmuer, og det for pattedyr mv. dødeligt giftige cicutoxin i Gifttyde og coniin i Skarntyde. Stofferne tåles af få insekter, men frastøder de fleste, som ellers vil æde planterne. Planterne beskytter især værdifulde unge blade med en lang levetid foran sig samt frø og frugter, som skal føre arten videre.
Det skaber et våbenkapløb mellem plantens udvikling af nye gifte og dyrenes evne til at tåle eller neutralisere giftstofferne; det antages at føre til en gensidig specialisering (kaldet co-evolution) og tilpasning mellem planter og dyr, som fremmer dannelsen af nye arter. Selv nyder vi smagen af kanel, kommen, peber, sennep og chili i passende doser, men stofferne er altså udviklet som forsvar mod insekter eller andre dyr, som vil æde planterne.
Om forfatterne


Kaj Sand-Jensen er professor i ferskvands-økologi ved Biologisk Institut, Københavns Universitet.
ksandjensen@bio.ku.dk
Jens Christian Schou er botaniker, forfatter og pensioneret lærer.
jcschou@gmail.com